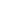多次跨膜靶点抗体开发难?用NovoiSMART®技术解决
使用GPRC5A的胞外序列对GPRC5D的4个胞外结构域 ECD进行突变替换,并命名1-4(对应图2结构的Loop1-4的替换)。结果如图3和图4显示,成功筛选出表位多样的GPRC5D候选分子。基于此类,对于部分耐药靶标,NovoiSMART®技术平台可以为您提供更优的选择! 图3.流式检测相较野生型的MFI下降比例热图 图4.🆚开云·Kaiyun网页版GPRC5D抗体表位多样性统计 二、CLDN6高特异性抗体筛选 Claudin6 (CLDN6) 是一个四次跨膜蛋白,具有一个短的细胞质N端和一个C端。

14岁上清华,29岁获普林斯顿终身教职,王梦迪最新论文,开发能读懂mRNA的语言模型,助力mRNA疫苗设计
自然界中的生命都遵循一个基本法则——中心法则,即细胞生命的遗传信息从DNA向RNA再向蛋白质的流动,DNA储存了遗传信息,蛋白组负责了细胞的结构和功能,而mRNA将DNA中的遗传信息翻译为蛋白组。实际上,mRNA中只有一部分会被翻译为蛋白质,其余部分是非翻译区,发挥着调控翻译的功能。5'非翻译区(5 ' UTR) 是mRNA序列开头的一段区域,位于蛋白质编码序列之前。它影响着mRNA分子的稳定性、定位和翻译,在调控mRNA到蛋白质的翻译过程中发挥🈺着至关重要的作用。
爱科瑞思、非同生物、英百瑞生物等在开发,肿瘤免疫新靶点5T4有何潜力?
该蛋白在胞外结构域中包含一个细胞外区、一个跨膜区(356-376aa)和一个胞内区(如下图所示)[2,3],其中胞内区含有PDZ结合基序(Ser-Asp-Va🍆开云·Kaiyun网页版l),该结合基序可以与位于细胞附近囊泡相关的细胞质蛋白膜TIP-2/GIPC互相作用,从而可参与细胞迁移和黏附信号转导过程的调控[4]。▲5T4结构(截图来源:Front Mol Neurosci [5]) 研究发现,5T4广泛表达于胚胎发育期的各种滋养层细胞,其在成人正常组织中表达受限,只在几种上皮细胞中表达,但却在很。
研究揭示连接组蛋白H5介导的核小体结合与染色质折叠和高级结构形成机制
迄今为止,只有占全长连接组蛋白的1/3左右的GD结构域的结构得到解析。该研究获得了连接组蛋白H5的全长蛋白,包括其N端结构域NTD和C端结构域CTD的原子分辨率结构,建立了完整染色质小体的原子分辨率结构。研究显示,在H5-30nm染色质纤维中,连接组蛋白H5以非对称三点接触方式结合在核小体核心颗粒上。其中,H5的CTD结构域形成经典的HMG-like螺旋折叠方式以紧密结合在linker DNA上,并通过H5的NTD和CTD结构域与连接DNA的相互作用促进染色质高级结构的折叠和。
Nature | 轰动有机合成界,寒门博士继Nature发文后,最新成果又登顶Cell
测序数据质量控制和检查数据比对和多匹配问题计算结合峰位置IGV中组学结果可视化差异peaks分析结合程度矩阵计算富集热图和曲线图绘制第五天理论部分机器学习概述线性模型决策树支持向量机集成学习模型选择与性能优化实操内容决策树算法实现随机森林算法实现支持向量机(SVM)算法实现朴素贝叶斯算法实现Xgboost算法实现聚类算法实现DBSCAN算法实现层次聚类算法实现理论部分基因功能注释和富集分析WGCNA(Weighted Gene Co-💥expression Network An。